Analyzing the Response Behavior of Lumbriculus variegatus (Oligochaeta: Lumbriculidae) to Different Concentrations of Copper Sulfate Based on Line Body Shape Detection and a Recurrent Self-Organizing Map

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Organisms and Observation System
2.2. Detection of Line Movement
2.3. Analysis of Movement Data
- Speed (mm/s): Average of movement distance of the test organisms in 1 min;
- Stop duration (s): Average time during which the test specimen did not move;
- Stop number: Stop frequency during 1 min;
- Turning rate (rad/s): The sum of angular changes in radians in absolute values divided by the cumulated time duration of movement;
- Meander (rad/s): The sum of angular changes in radians in absolute values divided by the cumulated movement distance.
3. Results
3.1. Effects on Body Length and Parameters
3.2. Effects on Angles of Body Segments
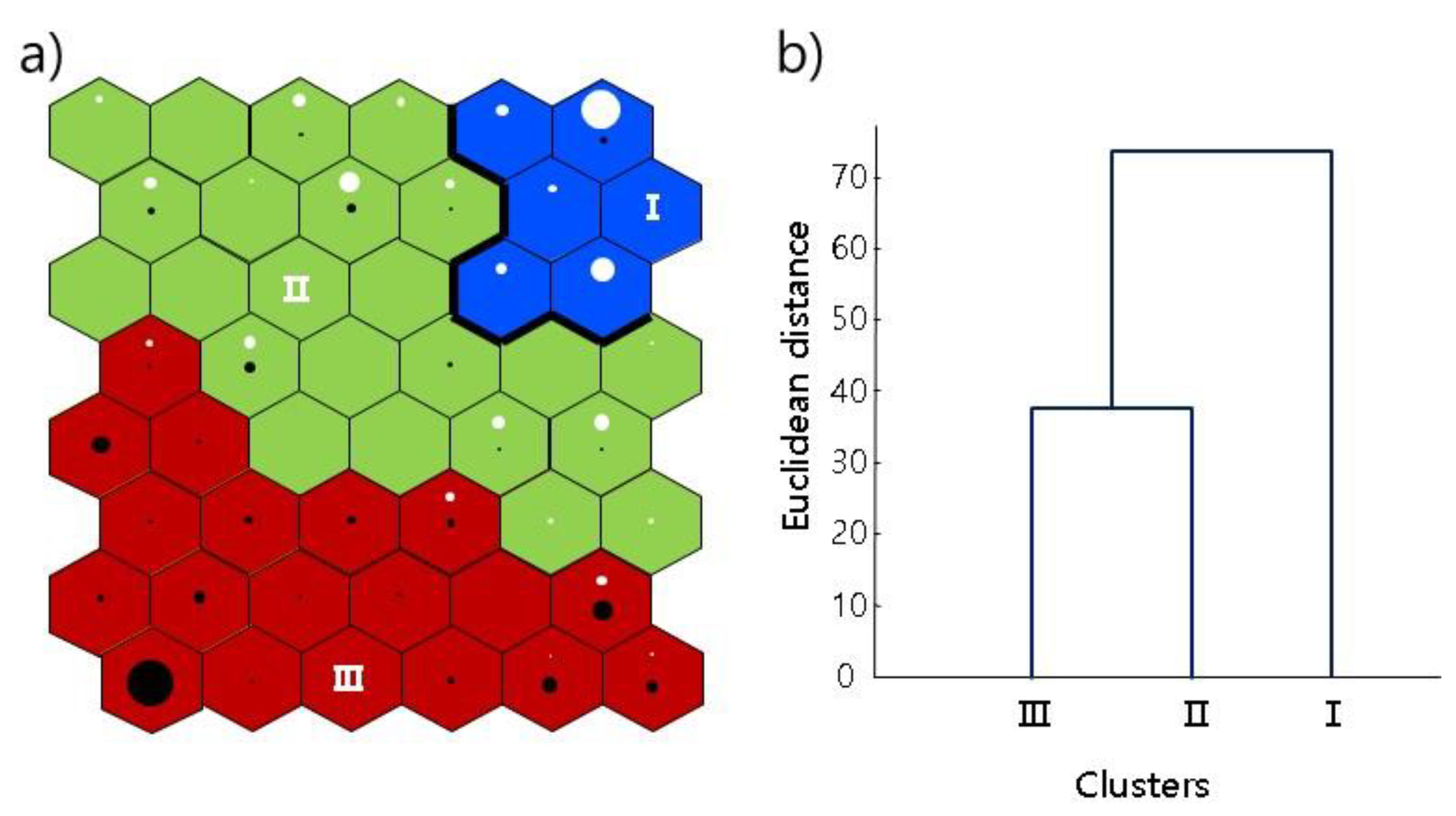
3.3. Patterning Sequential Line Movement
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lemly, A.D.; Smith, R.J. A behavioral assay for assessing effects of pollutants of fish chemoreception. Ecotoxicol. Environ. Saf. 1986, 11, 210–218. [Google Scholar] [CrossRef]
- Gerhardt, A.; Carlsson, A.; Ressemann, C.; Stich, K.P. New online biomonitoring system for Gammarus pulex (L.) (Crustacea): In situ test below a copper effluent in south Sweden. Environ. Sci. Technol. 1998, 32, 150–156. [Google Scholar] [CrossRef]
- Teather, K.; Harris, M.; Boswell, J.; Gray, M. Effects of Acrobat MZ® and Tattoo C® on Japanese medaka (Oryzias latipes) development and adult male behavior. Aquat. Toxicol. 2001, 51, 419–430. [Google Scholar] [CrossRef]
- Borcherding, J. Ten years of practical experience with the Dreissena-Monitor, a biological early warning system for continuous water quality monitoring. Hydrobiologia 2006, 556, 417–426. [Google Scholar] [CrossRef]
- Ji, C.W.; Lee, S.H.; Choi, K.-H.; Kwak, I.-S.; Lee, S.G.; Cha, E.Y.; Lee, S.-K.; Chon, T.-S. Monitoring of movement behaviors of chironomid larvae after exposure to diazinon by using fractal dimension and Self-Organizing Map. Int. J. Ecodyn. 2007, 2, 27–38. [Google Scholar] [CrossRef]
- Bae, M.-J.; Park, Y.-S. Biological early warning system based on the responses of aquatic organisms to disturbances: A review. Sci. Total Environ. 2014, 466–467, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Dutta, H.; Marcelino, J.; Richmonds, C. Brain acetylcholinesterase activity and optomotor behavior in bluegills, Lepomis macrochirus exposed to different concentrations of diazinon. Arch. Int. Physiol. Biochim. Biophys. 1992, 100, 331–334. [Google Scholar] [PubMed]
- Chon, T.-S.; Park, Y.-S.; Ross, M.H. Activity of German cockroach, Blattella germanica (L.) (Orthoptera : Blattellidae), at different microhabitats in semi-natural conditions when treated with sublethal doses of pesticides. J. Asia-Pac. Entomol. 1998, 1, 99–107. [Google Scholar]
- Chon, T.-S.; Park, Y.-S.; Park, J.Y.; Choi, S.-Y.; Kim, K.T.; Cho, E.C. Implementation of computational methods to pattern recognition of movement behavior of Blattella germanica (Blattaria : Blattellidae) treated with Ca2+ signal inducing chemicals. Appl. Entomol. Zool. 2004, 39, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Roast, S.D.; Widdows, J.; Jones, M.B. Disruption of swimming in the hyperbenthic mysid Neomysis integer (Peracarida: Mysidacea) by the organophosphate pesticide chlorpyrifos. Aquat. Toxicol. 2000, 47, 227–241. [Google Scholar] [CrossRef]
- Ibrahim, W.L.F.; Furu, P.; Ibrahim, A.M.; Christensen, N.Ø. Effect of the organophosphorous insecticide, chlorpyrifos (Dursban), on growth, fecundity and mortality of Biomphalaria alexandrina and on the production of Schistosoma mansoni cercariae in the snail. J. Helminthol. 1992, 66, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.A.; Teather, K.L.; Metcalfe, C.D. Reproductive success and behavior of Japanese Medaka (Oryzias latipes) exposed to 4-tert-octylphenol. Environ. Toxicol. Chem. 1999, 18, 2587–2594. [Google Scholar]
- Park, Y.-S.; Chung, N.-I.; Choi, K.-H.; Cha, E.Y.; Lee, S.-K.; Chon, T.-S. Computational characterization of behavioral response of medaka (Oryzias latipes) treated with diazinon. Aquat. Toxicol. 2005, 71, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.W.; Lee, S.H.; Kwak, I.-S.; Cha, E.Y.; Lee, S.-K.; Chon, T.-S. Computational analysis of movement behaviors of medaka (Oryzias latipes) after the treatments of copper by using fractal dimension and artificial neural networks. In Environmental Toxicology; Kungolos, A.G., Brebbia, C.A., Samaras, C.P., Popov, V., Eds.; WIT Press: Southampton and Mykonos, UK, 2006; pp. 93–107. [Google Scholar]
- Kwak, I.-S.; Chon, T.-S.; Kang, H.-M.; Chung, N.-I.; Kim, J.-S.; Koh, S.C.; Lee, S.-K.; Kim, Y.-S. Pattern recognition of the movement tracks of medaka (Oryzias latipes) in response to sub-lethal treatments of an insecticide by using artificial neural networks. Environ. Pollut. 2002, 120, 671–681. [Google Scholar] [CrossRef]
- Gerhardt, A. Importance of exposure route for behavioural responses in Lumbriculus variegatus Müller (Oligochaeta: Lumbriculida) in short-term exposures to Pb. Environ. Sci. Pollut. Res. 2007, 14, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.A.; Gabel, C.V.; Gabel, H.; Samuel, A.D.T. Temporal activity patterns in thermosensory neurons of freely moving Caenorhabditis elegans encode spatial thermal gradients. J. Neurosci. 2007, 27, 6083–6090. [Google Scholar] [CrossRef]
- Baek, J.-H.; Cosman, P.; Feng, Z.; Silver, J.; Schafer, W.R. Using machine vision to analyze and classify Caenorhabditis elegans behavioral phenotypes quantitatively. J. Neurosci. Methods 2002, 118, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Geng, W.; Cosman, P.; Berry, C.C.; Feng, Z.; Schafer, W.R. Automatic tracking, feature extraction and classification of C. elegans phenotypes. IEEE Trans. Biomed. Eng. 2004, 51, 1811–1820. [Google Scholar] [CrossRef]
- Stephens, G.J.; Johnson-Kerner, B.; Bialek, W.; Ryu, W.S. Dimensionality and dynamics in the behavior of C. elegans. PLoS Comput. Biol. 2008, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Hardaker, L.A.; Singer, E.; Kerr, R.; Zhou, G.; Schafer, W.R. Serotonin modulates locomotory behavior and coordinates egg-laying and movement in Caenorhabditis elegans. J. Neurobiol. 2001, 49, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Cronin, C.J.; Wittig, J.H.J.; Sternberg, P.W.; Schafer, W.R. An imaging system for standardized quantitative analysis of C. elegans behavior. BMC Bioinform. 2004, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ajie, B.C.; Estes, S.; Lynch, M.; Phillips, P.C. Behavioral degradation under mutation accumulation in Caenorhabditis elegans. Genetics 2005, 170, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, H.-J.; Kim, H.; Kim, B.-M.; Chon, T.-S.; Choi, J. Integrative assessment of benzene exposure to Caenorhabditis elegans using computational behavior and toxicogenomic analyses. Environ. Sci. Technol. 2014, 48, 8143–8151. [Google Scholar] [CrossRef] [PubMed]
- Bilbao, A.; Patel, A.K.; Rahman, M.; Vanapalli, S.A.; Blawzdziewicz, J. Roll maneuvers are essential for active reorientation of Caenorhabditis elegans in 3D media. Proc. Natl. Acad. Sci. USA 2018, 115, E3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awual, M.R. New type mesoporous conjugate material for selective optical copper(II) ions monitoring & removal from polluted waters. Chem. Eng. J. 2017, 307, 85–94. [Google Scholar] [CrossRef]
- Awual, M.R.; Hasan, M.M.; Rahman, M.M.; Asiri, A.M. Novel composite material for selective copper(II) detection and removal from aqueous media. J. Mol. Liq. 2019, 283, 772–780. [Google Scholar] [CrossRef]
- Awual, M.R.; Hasan, M.M.; Khaleque, M.A.; Sheikh, M.C. Treatment of copper(II) containing wastewater by a newly developed ligand based facial conjugate materials. Chem. Eng. J. 2016, 288, 368–376. [Google Scholar] [CrossRef]
- Petersen, E.J.; Huang, Q.; Weber, W.J. Ecological uptake and depuration of carbon nanotubes by Lumbriculus variegatus. Environ. Health Perspect. 2008, 116, 496–500. [Google Scholar] [CrossRef] [Green Version]
- US EPA. Methods for Measuring the Toxicity and Bioaccumulation of Sediment-associated Contaminants with Freshwater Invertebrates; U.S. Environmental Protection Agency: Washington, DC, USA, 2000.
- Phipps, G.L.; Ankley, G.T.; Benoit, D.A.; Mattson, V.R. Use of the aquatic oligochaete Lumbriculus varieatus for assessing the toxicity and bioaccumulation of sediment-associated contaminants. Environ. Toxicol. Chem. 1993, 12, 269–279. [Google Scholar] [CrossRef]
- O’Gara, B.A.; Bohannon, V.K.; Teague, M.W.; Smeaton, M.B. Copper-induced changes in locomotor behaviors and neuronal physiology of the freshwater Oligochaete, Lumbriculus variegatus. Aquat. Toxicol. 2004, 69, 51–66. [Google Scholar] [CrossRef]
- Drewes, C.D. Helical swimming and body reversal behaviors in Lumbriculus variegatus (Annelida: Clitellata: Lumbriculidae). Hydrobiologia 1999, 406, 263–269. [Google Scholar] [CrossRef]
- Gerhardt, A.; de Bisthoven, L.J.; Penders, E. Quality control of drinking water from the River Rhine with the multispecies freshwater biomonitor. Aquat. Ecosyst. Health Manag. 2003, 6, 159–166. [Google Scholar] [CrossRef]
- Kim, C.-K.; Kwak, I.-S.; Cha, E.-Y.; Chon, T.-S. Implementation of wavelets and artificial neural networks to detection of toxic response behavior of chironomids (Chironomidae: Diptera) for water quality monitoring. Ecol. Model. 2006, 195, 61–71. [Google Scholar] [CrossRef]
- Son, K.H.; Ji, C.W.; Park, Y.-M.; Cui, Y.; Wang, H.Z.; Chon, T.-S.; Cha, E.Y. Recurrent Self-Organizing Map implemented to detection of temporal line-movement patterns of Lumbriculus variegates (Oligochaeta: Lumbriculidae) in response to the treatments of heavy metal. In Environmental Toxicology; Kungolos, A.G., Brebbia, C.A., Samaras, C.P., Popov, V., Eds.; WIT Press: Southampton/Mykonos, UK, 2006; pp. 93–107. [Google Scholar]
- Ji, C.W.; Kim, H.; Park, Y.-S.; Chon, T.-S. Computational analysis of movement behaviors of medaka (Oryzias latipes) in response to chemical and thermal stressors. J. Korean Phys. Soc. 2012, 60, 570–575. [Google Scholar] [CrossRef]
- Gershenfeld, N.; Weigend, A. The future of time series: Learning and understanding. In Time Series Prediction: Forecasting the Future and Understanding the Past; Weigend, A., Gershnfeld, N., Eds.; Addison-Wesley: Reading, MA, USA, 1993; pp. 1–70. [Google Scholar]
- Chon, T.S.; Park, Y.S.; Cha, E.Y. Patterning of community changes in benthic macroinvertebrates collected from urbanized streams for the short time prediction by temporal artificial neuronal networks. In Artificial Neuronal Networks; Lek, S., Guégan, J.-F., Eds.; Springer: Berlin/Heidelberg, Germany, 2000; pp. 99–114. [Google Scholar] [CrossRef]
- Chon, T.-S.; Kwak, I.-S.; Park, Y.-S.; Kim, T.-H.; Kim, Y. Patterning and short-term predictions of benthic macroinvertebrate community dynamics by using a recurrent artificial neural network. Ecol. Model. 2001, 146, 181–193. [Google Scholar] [CrossRef]
- Kohonen, T. Self-Organizing Maps; Springer: Berlin, Germany, 2001. [Google Scholar]
- Varsta, M.; Heikkonen, J.; Millan, J.d.R. Context learning with the self organizing map. In Proceedings of the Proceedings of WSOM 1997, Workshop on Self-Organizing Maps, Espoo, Finland, 4–6 June 1997; pp. 197–202. [Google Scholar]
- Timo, K.; Varsta, M.; Heikkonen, J.; Kaski, K. Temporal sequence processing using recurrent SOM. In Proceedings of the 2nd International Conference on Knowledge-Based Intelligent Engineering Systmes, Adelaide, Australia, 21 April 1998; pp. 290–297. [Google Scholar]
- Angelovič, P. Time series prediction using RSOM and local models. In Proceedings of the IIT. SRC 2005 Student Research Conference, Bratislava, Slovakia, 27 May 2005; pp. 27–34. [Google Scholar]
- Bailey, H.C.; Liu, D.H.W. Lumbriculus variegatus, a benthic Oligochaete, as a bioassay organism. In Aquatic Toxicology Hazard Assessment 3rd Symposium, ASTM STP 707; Eaton, J.C., Parrish, P.R., Hendricks, A.C., Eds.; ASTM: Philadelphia, PA, USA, 1980. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–656. [Google Scholar] [CrossRef] [Green Version]
- StatSoft Inc. STATISTICA (Data Analysis Software System), Version 7; StatSoft Inc.: Tulsa, Oklahoma, 2004. [Google Scholar]
- The Mathworks. MATLAB, Version 7.0; The Mathworks, Inc.: Massachusetts, MA, USA, 2001. [Google Scholar]
- Mielke, E.W.; Berry, K.J.; Johnson, E.S. Multiresponse permutation procedures for a priori classifications. Commun. Stat. Theory Methods 1976, 5, 1409–1424. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. PC-ORD. Multivariate Analysis of Ecological Data. Version 4.41; MjM Software: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Lesiuk, N.M.; Drewes, C.D. Autotomy reflex in a freshwater oligochaete, Lumbriculus variegatus (Clitellata: Lumbriculidae). Hydrobiologia 1999, 406, 253–261. [Google Scholar] [CrossRef]
- ASTM (American Society for Testing and Materials). Annual Book of ASTM Standards. Volume 11.05; ASTM: Philadelphia, PA, USA, 1995. [Google Scholar]
- Rogge, R.W.; Drewes, C.D. Assessing sublethal neurotoxicity effects in the freshwater oligochaete, Lumbriculus variegatus. Aquat. Toxicol. 1993, 26, 73–90. [Google Scholar] [CrossRef]
- Ding, J.; Drewes, C.D.; Hsu, W.H. Behavioral effects of ivermectin in a freshwater oligochaete, Lumbriculus variegatus. Environ. Toxicol. Chem. 2001, 20, 1584–1590. [Google Scholar] [CrossRef]
- Chapman, K.K.; Benton, M.J.; Brinkhurst, R.O.; Scheuerman, P.R. Use of the aquatic oligochaetes Lumbriculus variegatus and Tubifex tubifex for assessing the toxicity of copper and cadmium in a spiked-artificial-sediment toxicity test. Environ. Toxicol. 1998, 14, 271–278. [Google Scholar] [CrossRef]
- GERBE. Programme de Recherche: Toxicologie - Ecotoxicologie des Pesticides et des Metaux Lourds. Deuxieme Rapport d’Activites; Laboratory of Eco-Toxicology, Faculty of Sciences, University of Reims Champagne: Ardenne, France, 1998. [Google Scholar]
- Awual, M.R. Novel ligand functionalized composite material for efficient copper(II) capturing from wastewater sample. Compos. Part B Eng. 2019, 172, 387–396. [Google Scholar] [CrossRef]
- Awual, M.R. A novel facial composite adsorbent for enhanced copper(II) detection and removal from wastewater. Chem. Eng. J. 2015, 266, 368–375. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Speed | Stop Duration | Stop Number | Turning Rate | Meander |
|---|---|---|---|---|---|
| p1 | n.s *. | n.s. | n.s. |  | n.s. |
| p2 | a | n.s. | n.s. | | n.s. |
| p3 | a | n.s. | n.s. | | n.s. |
| p4 | a | n.s. | n.s. | | n.s. |
| p5 | a | n.s. | n.s. | | n.s. |
| p6 | a | n.s. | n.s. | | n.s. |
| p7 | a | n.s. | n.s. | | n.s. |
| p8 | a | n.s. | n.s. | | n.s. |
| p9 | a | n.s. | n.s. | | n.s. |
| p10 | a | n.s. |  | | n.s. |
| p11 | a | n.s. | |  | n.s. |
| p12 | a | n.s. | | | n.s. |
| p13 | a | n.s. | | | n.s. |
| a1 | a2 | a3 | a4 | a5 | ||
| F | 100.89 | 35.16 | 25.16 | 10.60 | 5.09 | |
| P | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0056 | |
| Group | I ≠ II ≠ III | I ≠ II ≠ III | I ≠ II ≠ III | I ≠ II = III | I ≠ II = III | |
| a6 | a7 | a8 | a9 | a10 | a11 | |
| F | 33.82 | 22.26 | 21.03 | 40.61 | 55.64 | 70.96 |
| P | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Group | I ≠ II = III | I ≠ II = III | I ≠ II = III | I ≠ II = III | I ≠ II ≠ III | I ≠ II ≠ III |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, C.W.; Park, Y.-S.; Cui, Y.; Wang, H.; Kwak, I.-S.; Chon, T.-S. Analyzing the Response Behavior of Lumbriculus variegatus (Oligochaeta: Lumbriculidae) to Different Concentrations of Copper Sulfate Based on Line Body Shape Detection and a Recurrent Self-Organizing Map. Int. J. Environ. Res. Public Health 2020, 17, 2627. https://doi.org/10.3390/ijerph17082627
Ji CW, Park Y-S, Cui Y, Wang H, Kwak I-S, Chon T-S. Analyzing the Response Behavior of Lumbriculus variegatus (Oligochaeta: Lumbriculidae) to Different Concentrations of Copper Sulfate Based on Line Body Shape Detection and a Recurrent Self-Organizing Map. International Journal of Environmental Research and Public Health. 2020; 17(8):2627. https://doi.org/10.3390/ijerph17082627
Chicago/Turabian StyleJi, Chang Woo, Young-Seuk Park, Yongde Cui, Hongzhu Wang, Ihn-Sil Kwak, and Tae-Soo Chon. 2020. "Analyzing the Response Behavior of Lumbriculus variegatus (Oligochaeta: Lumbriculidae) to Different Concentrations of Copper Sulfate Based on Line Body Shape Detection and a Recurrent Self-Organizing Map" International Journal of Environmental Research and Public Health 17, no. 8: 2627. https://doi.org/10.3390/ijerph17082627