
Cold Storage Period of Bulbs Influences Their Sugar Contents and Post-Storage Growth and Flowering in Lilium Oriental Hybrids


Department of Horticultur and Life Science, Yeungnam University, Gyeongsan 38541, Korea
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(11), 1080; https://doi.org/10.3390/agriculture11111080
Submission received: 23 September 2021
/
Revised: 28 October 2021
/
Accepted: 30 October 2021
/
Published: 1 November 2021
(This article belongs to the Section Crop Production)
Abstract
:This study was carried out to investigate the changes in internal shoot growth, total and reducing sugar content, plant growth, and flowering characteristics after cold-stored bulbs of Lilium Oriental Hybrids are planted. Bulbs of Lilium Oriental Hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’ were harvested at the end of November, cleaned, treated with fungicides, conditioned for 30 days at 4 °C, and cold-stored for 30 to 120 days at 0 °C. Plant height, fresh and dry weights, leaf numbers and area, chlorophyll content (SPAD index), days to flowering, number of flower buds, and flower diameter were measured when the cut flowers were harvested. In all cultivars, the shoot lengths in the bulbs increased as the storage period increased. As the storage period was extended, the fructose and glucose contents increased, whereas sucrose content decreased. The total sugar content increased during the early storage period and decreased subsequently. Plant height, fresh and dry weights of cut flowers showed a decreasing trend as the storage period increased. Flower diameter generally decreased with increasing storage period but varied among cultivars. These results indicated that more storage carbohydrates in bulbs were consumed as the storage period increased, and thus, the subsequent plant growth and flowering were negatively affected.
1. Introduction
The quality of lily bulbs is known to be affected considerably by the conditions of plant growth and subsequent storage. During the growth of Lilium rubellum, flower bud initiation in the bulbs occurs in mid-September and is completed through November and December [1,2]. During the cold storage of bulbs, the sucrose content increases by the hydrolysis of starch in the bulb [3,4]. For example, an increase in the sucrose and reducing sugars was observed in L. rubellum [5], similar to that reported in other species such as Easter lily [6], hyacinth, and tulip [7].
The lily bulb is dormant after harvest and cold treatment in a forcing culture is commonly needed to break dormancy. Cold treatment is necessary to induce vernalization [8], and the Asiatic and Oriental Hybrids lilies must be stored at 1.7 °C for 6 to 8 weeks [9].
Lilium Oriental Hybrids are very important cut-flower crops worldwide, so improvement in yield and quality, in addition to diversification of the production period, are important issues in the lily industry. To meet the demands for the efficient production of cut flowers, a detailed understanding of the physiological changes in lily bulbs based on storage conditions and period is needed. Bulbs grown domestically are expected to be physiologically different from imported bulbs.
There is limited information on the effects of storage period and conditions on bulb quality and subsequent plant growth in Lilium Oriental Hybrids grown in South Korea. To ensure the supply of high-quality bulbs grown under Korean field conditions, it is necessary to elucidate the physiological and morphological changes in the bulb under cold storage and optimal storage periods. This study was performed to investigate the effects of storage period on the morphological characteristics and carbohydrate content of the bulbs, and post-storage growth and flowering in Oriental Hybrid lilies.
2. Materials and Methods
2.1. Plant Materials
Bulbs of Lilium Oriental hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’ grown and harvested from the open field of a lily export co-op in Taean, Korea (36°71′09.3″ N, 126°31′76.3″ E) were used in this experiment. Bulbs selected based on size (≥10 cm in circumference) were planted in February, 2017, cultivated in the open field using standard cultural practices [10], and harvested in the last week of November. After harvest, the bulbs were graded for uniform size (16–18 cm in circumference), and the stem and root were removed. The bulbs were dried in the air for 2 h after washing the surface soil with running water and wiping off the surface water.
After the bulbs were measured, they were treated with a mixture solution of insecticides and fungicides (active ingredients: dimethoate, diazinon, captan, and carbosulfan) for 30 min, air-dried, and placed in cold storage. For cold storage treatment, the bulbs were conditioned for 30 days at 4 °C and stored at 0 °C for 30–120 days at 10-day intervals. After the designated storage periods, the bulbs were conditioned at 12 °C for 2 weeks and planted in a greenhouse for growth and flowering. Therefore, the bulbs cold-stored for 30 days were firstly planted from 13 February, 2018 after pre-conditioning for 30 days, cold storage for 30 days, and post-conditioning for 2 weeks.
2.2. Measurements of Morphological Characteristics and Total and Free Sugars of Bulbs at the End of Each Storage Period
The length of the new shoots in the bulb were measured after removing the surrounding scales. The length was measured from the basal plate to the end of the shoot.
Total sugar content was measured using the phenol-sulfuric acid method [11]. The bulb was cut in half longitudinally, and the base was discarded. The scale tissue was sliced into 0.5-cm thickness and 5-g sample was taken. The sample was macerated in a mortar containing 50 mL of distilled water. The homogenate was centrifuged at 8000 rpm for 20 min. The supernatant was filtered through Whatman No. 1 filter paper (GE Healthcare, Amersham, UK), the volume was adjusted to 50 mL, and it was stored at −20 °C until analysis. The total sugar content was calculated using a standard curve (y = 0.0082x × 0.0064).
The free sugar content was measured using high-performance liquid chromatography (HPLC). Free sugar samples were extracted from the tissues as described for the total sugar sample. The 5-g sample was placed in a 250-mL flask, to which 50 mL of ethanol was added. The mixture was heated to 65 °C in a water bath for 1 h using a reflux cooling system. The extract was filtered through a 0.45-µm syringe filter, and 10 µL of the sample was injected into an HPLC system for analysis (RI-2031 Plus, Jasco, Tokyo, Japan). A carbohydrate column (WAT084038, 4.6 mm × 250 mm, 5 µm, Waters, Milford, MA, USA) was used for separation using 74% acetonitrile as the mobile phase at a flow rate of 1 mL/min. The column temperature was maintained at 38 °C, and an RI detector was used for the detection.
2.3. Measurements of Plant Growth and Flowering Characteristics
As described in the preceding sections, the lily bulbs were conditioned and stored for 30 to 120 days at 0 °C and planted for regrowth and flowering. Planting boxes (L 64.3 cm × W 24.0 cm × H 17.5 cm) were filled with a medium made of coco-peat and perlite (14:1, v/v) thoroughly mixed with 36 g of granule fertilizer (Osmocote Plus 15-11-13 + 2 MgO + TE, ICL Specialty Fertilizers, Geldermalsen, The Netherlands). Four bulbs were planted per box and were allowed to grow at 25/20 °C (day/night) under 20‒25 μmol∙m−2∙s−1 PPFD light conditions for 2 weeks and transferred to the greenhouse in the affiliated farm of Yeungnam University, Gyeongsan, Korea (35°49′38.1′′ N, 128°45′20.0′′ E). The plants were irrigated once per week with tap water. The greenhouse environment was maintained at 23/17 °C (day/night) in February and April, 26/21 °C in May and June, and 28/23 °C in July and August. The temperatures of the media were 18/22 °C (min/max) in February and April, 23/27 °C in May and June, and 24/28 °C in July and August. The light intensity was controlled using 35% and 65% shading cloths throughout the experiment.
The entire plant was harvested when the first flower was completely opened, and plant height, stem diameter, leaf number, fresh weight, and dry weight of the plant were measured. For dry weight measurement, the cut flowers harvested were placed in an oven set to 80 °C for 3 days, and the weight was measured. Days to flowering, number of unopened flowers, and petal width were recorded. Chlorophyll content was estimated as SPAD index using a chlorophyll meter (SPAD-502, Minolta) and leaf areas using a Win DIAS 3 Image analysis system (Delta-T Devices, Cambridge, UK).
2.4. Statistical Analysis of Collected Data
Each treatment consisted of three replicates with twelve bulbs or plants. Statistical analysis and treatment means were separated via Tukey’s honestly significant difference test using SAS software (SAS 9.1.3, SAS Institute, Cary, NC, USA). Regression analysis was performed using the SigmaPlot V10 program (SPSS Software, Chicago, IL, USA).
3. Results and Discussion
3.1. Shoot Length and Carbohydrates Contents of Bulbs According to Storage Period
The shoot length in bulbs measured at the end of cold storage increased with increasing storage period for all three cultivars (Figure 1). In ‘Le Reve’, the shoot length in the bulb after 30 days of cold storage was 21.4 mm, and continuously increased to 48.5 mm after 120-day storage (Figure 1a). Likewise, the shoot lengths after 30- to 120-day storage increased from 25.8 to 43.3 mm in ‘Sorbonne’ and from 23.1 to 36.0 mm in ‘Siberia’ (Figure 1b,c). In addition, there were statistically significant correlations between storage period and shoot length in ‘Le Reve’ (y = 0.004x2 − 0.34x + 29.2, R2 = 0.97, p < 0.001), ‘Sorbonne’ (y = 0.003x2 − 0.35x + 34.6, R2 = 0.92, p < 0.001), and ‘Siberia’ (y = 0.002x2 − 0.25x + 29.1, R2 = 0.86, p < 0.001).
Sugar content in bulbs at the end of cold storage was influenced by the storage period (Figure 2). Fructose content in bulbs increased in ‘Le Reve’ and ‘Siberia’ as the storage period increased (Figure 2a,c). In ‘Le Reve’, fructose content has increased from 8.3 mg·g−1 at 30 days to 18.85 mg·g−1 at 120 days in storage. In ‘Siberia’, the content was increased from 3.85 to 14.05 mg·g−1 at 110 days. In ‘Sorbonne’, there was a variation in fructose content during the storage period, but the trend indicated an increase in fructose content from ~10 to ~20 mg·g−1 at 120 days (Figure 2b). Our results showed that there was an approximate increase of 10 mg·g−1 in fructose content during the storage period.
The glucose content showed a trend similar to that of fructose, with an increasing trend as the storage period was extended (Figure 2d–f) but at a reduced scale. In ‘Le Reve’, the glucose content increased from 1.7 to 8.6 mg·g−1 as the storage period increased from 30 to 120 days (Figure 2d). In ‘Sorbonne’, there was a marked increase from 1.2 to 10.55 mg·g−1, while ‘Siberia’ exhibited an increase from 1.3 to 7.9 mg·g−1. Accumulation of glucose and fructose in lily bulbs was observed during the process of vernalization and flower bud differentiation because they are reducing sugars that can be used directly as respiratory and structural materials [12].
However, the sucrose content showed an opposite decreasing trend during storage (Figure 2g–i). The sucrose content in ‘Sorbonne’ was 38.65 mg·g−1 at 30 days, increased to 48.6 mg·g−1 at 70 days, and then decreased to 7.00 mg·g−1 at 120 days in storage. The sucrose content in ‘Siberia’ was 48.6 mg·g−1 at 30 days and steadily decreased to 20.85 mg·g−1 after 110 days of storage. Sucrose contents in ‘Le Reve’ showed a slightly decreasing trend during the storage period without statistical significance. In general, sucrose content was much higher than fructose and glucose content in these results. Changes in sucrose content during storage differed from those in glucose and fructose content because plant carbohydrates are mainly transported in the form of sucrose. That is, the mobile form of sucrose first increased and then decreased as the glucose and fructose content increased. These differences were in accordance with the finding by Ranwala and Miller [13] that the sucrose content was greater than that of reducing sugars.
The total sugar content increased during the early period, which then stabilized and remained unchanged thereafter (Figure 2j–l). In ‘Sorbonne’, the total sugar content was 614.4 mg·g−1 at 30 days in storage, gradually increased to 973.2 mg·g−1 at 100 days, and then decreased slightly to 824.6 mg·g−1. The contents in ‘Siberia’ showed a somewhat similar trend to ‘Sorbonne’, increasing from 668.4 mg·g−1 at 30 days to 937.4 mg·g−1 at 60 days, and reduced to 716.4 mg·g−1 at 120 days. In ‘Le Reve’, the change was not clear but showed an increasing trend throughout the storage period (Figure 2j). In a previous study, the total sugar content in Corydalis bracteata was not affected by the surrounding temperature [14]. In this study, the total sugar content did not change significantly, indicating that the total sugar content did not change with the storage period under constant storage temperature conditions. The internal shoots elongated continuously as the storage period was extended in all three cultivars. In general, it was observed that the glucose and fructose content increased, whereas the sucrose content decreased. Overall, the total carbohydrate content increased in the early period and then remained at a similar level or decreased slightly in the latter period.
Shin et al. (2002) reported that cold treatment induced metabolic activity in the lily scales, causing sprouting of the bulb and further plant development [15]. The sprouting process in the lily bulb induced by cold treatment is accompanied by the rapid hydrolysis of starch and accumulation of sucrose and reducing sugars [6,16]. Cold storage of the bulbs of Tulipa ‘Apeldoorn’ and Corydalis induces mobilization of nutrients from the storage tissues to the growing buds, and the movement was inhibited at high temperatures [13,17,18]. These findings and our results regarding shoot growth and changes in free and total sugar content indicate that the lily bulbs maintained the metabolic activity and supply of nutrition to the shoot during the cold storage period.
The increase and decrease in sucrose content in the bulb were reported to be related to the induction and breakage of dormancy, respectively [18,19,20,21]. Because storage starch is hydrolyzed and becomes a source of free sugars in geophyte bulbs during the fall and winter when photosynthesis cannot occur [22], we might estimate the degree of dormancy breakage by monitoring the sucrose content in the bulb.
3.2. Post-Storage Growth and Flowering
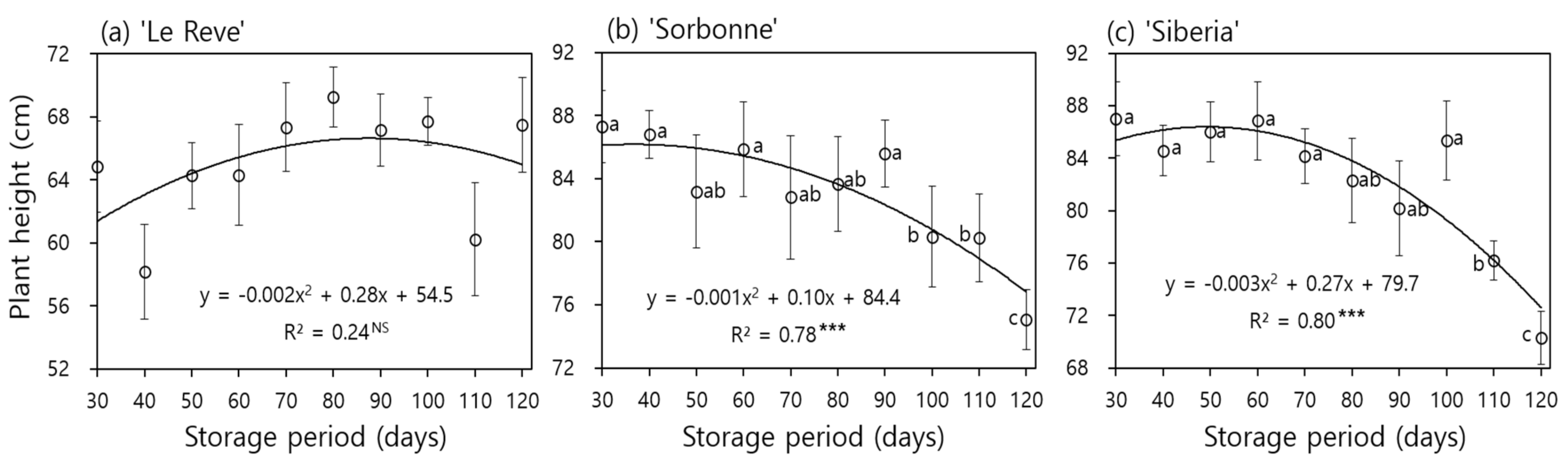
Plant heights after planting of bulbs in the greenhouse showed a decreasing trend as the storage period increased in ‘Sorbonne’ and ‘Siberia’ (Figure 3b,c). The plant height of ‘Sorbonne’ was 87.3 cm in 30-day stored bulbs and was reduced to 75.1 cm in 120-day storage (y = −0.001x2 + 0.10x + 84.4, R2 = 0.78, p < 0.001). Similarly, the plant height of ‘Siberia’ was 87.0 and 70.3 cm in bulbs stored for 30 and 120 days, respectively (y = −0.003x2 + 0.27x + 79.7, R2 = 0.80, p < 0.001). However, there was no significant difference among the mean values for each the cold storage periods from 30 to 90 days in ‘Sorbonne’ and from 30 to 100 days in ‘Siberia’. Yang et al. reported that plant height in ‘Sorbonne’ decreased with increasing cold-storage duration and the difference became significant after 60 to 70 days of storage [23]. The reason why long-term bulb storage reduced plant height is presumed to be due to the decrease in bulb reserves due to respiration and shoot growth during the storage period and the accelerated flowering under relatively higher temperature and longer daylength after bulb planting [22,23]. On the other hand, there was no clear reduction in plant height in ‘Le Reve’ and the height remained nearly the same, regardless of the storage period (Figure 3a).
Fresh weights of the cut flowers showed a clear decreasing trend in ‘Sorbonne’ and ‘Siberia’ with the increase in the storage period, but a tendency to increase and then decrease was observed in ‘Le Reve’ (Figure 4a–c). Fresh weight in ‘Sorbonne’ decreased from 161.7 g at 40 days to 82.1 g at 120 days (y = −0.003x2 + 0.002x + 127.4, R2 = 0.88, p < 0.001). Similarly, fresh weight in ‘Siberia’ decreased from 143.6 g at 30 days to 77.7 g at 120 days (y = −0.004x2 + 0.10x + 140.2, R2 = 0.64, p < 0.01). The dry weights of plants showed a very similar trend to that of the fresh weights, decreasing trends over the storage period (Figure 4d–f).
The plant height after cold storage decreased as the storage period was extended in ‘Sorbonne’ and ‘Siberia’ (Figure 3). The fresh and dry weights decreased similarly in the longer-stored bulbs (Figure 4). The reserves, such as carbohydrates, in Lilium bulbs are known to decrease as the stems and leaves grow after the bulb is planted, to be utilized in the growth and development of the stems and flowers [15,24,25]. These stored carbohydrates are also utilized as sources of metabolism during storage, and thus are decreased. Consequently, bulbs with longer storage periods had lower levels of carbohydrates than those with shorter storage periods and showed a reduced overall growth response in plant length and fresh and dry weights.
The number of leaves, leaf area, and chlorophyll content (SPAD index) differed according to storage period in all three cultivars (Table 1). In bulb plants growing at warmer temperatures, photosynthesis rates are accelerated, and starch accumulation is promoted and saturated; however, at lower temperatures, photosynthesis rates are reduced while starch accumulation is limited, and consequently leaf senescence is rapidly promoted [26]. In this study, the growing conditions after planting the bulbs in the greenhouse were changed in air and medium temperatures and day length. Therefore, there were significantly differences in leaf numbers, leaf area, and chlorophyll content among the storage periods (Table 1). However, these differences were of a negligible level in respect to commercial quality parameters.
Days to flowering decreased with increasing storage period in all three cultivars (Table 2) because bulbs had elongated shoots (Figure 1) and changed carbohydrate composition as storage period increased (Figure 2). In addition, they were grown under the environmental conditions such as higher temperature, more light quantity, and longer day length that can promote plant development as the planting date was delayed. There was a significant correlation between days to flowering and plant height and fresh and dry weight in ‘Sorbonne’ and ‘Siberia’ (Figure 5). The plant height and weight of lily cut flowers are greatly affected by photosynthesis during plant growth, as well as the size and nutrient content of the bulbs [27]. Therefore, it can be said that as the growth period before flowering increases, the amount of photosynthate increases, as does the plant height and weight.
The number of flower buds was not affected by storage period in the three cultivars (Table 2). The number of flower buds was reported to be influenced by the bulb size [28,29]. This was because the size of the flower meristem was also affected by the bulb size and weight [30,31]. However, in this study, we graded the bulb size and used similarly sized bulbs, and thus there seemed to be no difference in the number of flower buds, regardless of storage period.
Flower diameter showed different patterns according to storage period and cultivar (Table 2). There was a difference between early and later periods in ‘Le Reve’, the flower diameter was 16.6–17.1 cm between 30 and 60 days and was 15.1 cm after 70 days. There was no significant difference in the storage period in ‘Sorbonne’. In ‘Siberia’, the diameter was 23.8 cm at 40 days, decreased to 20.7 cm at 90 days, and remained similar afterwards.
Sucrose is the main sugar transported between tissues for growth in many plants [32]. Ranwala and Miller (1998) reported that high levels of sucrose are found in the flower bud tissue of Lilium longiflorum [33]. In our study, the sucrose content decreased as the storage period was extended (Figure 2). Since the flower diameter decreased as the storage period was extended, we presumed that the reduced sucrose content negatively affected flower diameter after the bulbs were replanted for flowering.
4. Conclusions
For the production of high-quality lily cut flowers, we believe that an optimum storage period with the minimal consumption of stored carbohydrates that ensures subsequent normal growth of plants and flowers after replanting must be determined. Our study indicated the optimum storage period to be 60 days for ‘Le Reve’ and 80‒90 days for ‘Sorbonne’ and ‘Siberia’. The short storage period in ‘Le Reve’ was probably due to the characteristics of the cultivar (i.e., early flowering), and mainly used for forced production; it showed a generally unstable production pattern after a long period of storage.
Author Contributions
J.K.: Methodology, investigation, visualization, software, and writing—original draft. W.O.: Conceptualization, methodology, project administration, supervision, funding acquisition, writing—review & editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Korea Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Smart Plant Farming Industry Technology Development Program, funded by Ministry of Agriculture, Food and Rural Affairs (MAFRA), Republic of Korea (grant number. 319067-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have influenced the work reported in this paper.
References
- Ikeda, Y. Studies on Lilium rubellum Baker. 2. Regarding variation and utilization as horticultural variety. Bull. Niigata Hortic. Exp. Stn. 1971, 6, 42–70. [Google Scholar]
- Niimi, Y.; Oda, M. Time of initiation and development of flower buds in Lilium rubellum Baker. Sci. Hortic. 1989, 39, 341–348. [Google Scholar] [CrossRef]
- Stitt, M.; Hurry, V. A plant for all seasons: Alterations in photosynthetic carbon metabolism during cold acclimation in Arabidopsis. Curr. Opin. Plant Biol. 2002, 5, 199–206. [Google Scholar] [CrossRef]
- Guy, C.; Kaplan, F.; Kopka, J.; Selbig, J.; Hincha, D.K. Metabolomics of temperature stress. Physiol. Plant. 2008, 132, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.Y.; Niimi, Y.; Han, D.S. Changes in endogenous abscisic acid and soluble sugars levels during dormancy-release in bulbs of Lilium rubellum. Sci. Hortic. 2006, 111, 68–72. [Google Scholar] [CrossRef]
- Miller, W.B.; Langhans, R.W. Low temperature alters carbohydrate metabolism in Easter lily bulbs. HortScience 1990, 25, 463–465. [Google Scholar] [CrossRef] [Green Version]
- Moe, R.; Wickstrøm, A.L.F. The effect of storage temperature on shoot growth, flowering, and carbohydrate metabolism in tulip bulbs. Physiol. Plant. 1973, 28, 81–87. [Google Scholar] [CrossRef]
- Miller, W.B. Lilium longiflorum. In The Physiology of Flower Bulbs; de Hertogh, A., Le Nard, M., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 391–422. [Google Scholar]
- Larson, R.A. Introduction to Floriculture, 2nd ed.; Academic Press Inc.: San Diega, CA, USA, 1992; pp. 195–212. [Google Scholar]
- Choi, S.T.; Ko, J.Y.; Paek, K.Y.; Kim, J.Y.; Kim, Y.J.; Shin, H.K. Lily: Cultivation Techniques of Bulbs and Cut flowers. The Farmers Newspaper: Seoul, Korea, 1996; pp. 137–196. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Gu, J.; Zeng, Z.; Wang, Y.; Lyu, Y. Transcriptome analysis of carbohydrate metabolism genes and molecular regulation of sucrose transport gene LoSUT on the flowering process of developing Oriental hybrid lily ‘Sorbonne’ bulb. Int. J. Mol. Sci. 2020, 21, 3092. [Google Scholar] [CrossRef]
- Ranwala, A.P.; Miller, W.B. Analysis of nonstructural carbohydrates in storage organs of 30 ornamental geophytes by high-performance anion-exchange chromatography with pulsed amperometric detection. New Phytol. 2008, 180, 421–433. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Miroslavov, E.A.; Shavarda, A.L.; Laberche, J.C.; Boitel-Conti, M. Bud development in corydalis (Corydalis bracteata) requires low temperature: A study of developmental and carbohydrate changes. Ann. Bot. 2010, 105, 891–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.S.; Chakrabarty, D.; Paek, K.Y. Sprouting rate, change of carbohydrate contents and related enzymes during cold treatment of lily bulblets regenerated in vitro. Sci. Hortic. 2002, 96, 195–204. [Google Scholar] [CrossRef]
- Matsuo, T.; Mizuno, T. Changes in the amount of two kinds of reserve glucose-containing polysaccharides during germination of the Easter lily bulb. Plant Cell Physiol. 1974, 15, 555–558. [Google Scholar]
- Khodorova, N.V. A Study of Adaptation to Cold in a Geophyte Species [Corydalis bracteata (Steph.) Pers, Fumariaceae DC.] and an Approach of Secondary Metabolism during Plant Development. Ph.D. Thesis, Jules Verne University of Picardy, Amiens, France, 2011. [Google Scholar]
- Hobson, G.E.; Davies, J.N. Mitochondrial activity and carbohydrate levels in tulip bulbs in relation to cold treatment. J. Exp. Bot. 1977, 28, 559–568. [Google Scholar] [CrossRef]
- Nowak, J.; Saniewsky, M.; Rudnicki, R.M. Studies on the physiology of hyacinth bulbs (Hyacinthus orientalis). I. Sugar content and metabolic activities in bulbs exposed to low temperatures. J. Hortic. Sci. 1974, 49, 383–390. [Google Scholar] [CrossRef]
- Hobson, G.E.; Davies, J.N. Influence of the extent and period of cold treatment on the flowering behaviour, composition and metabolic activity of tulip bulbs. Sci. Hortic. 1978, 8, 279–287. [Google Scholar] [CrossRef]
- Aguettaz, P.; Paffen, A.; Delvallée, I.; van der Linde, P.; de Klerk, G.J. The development of dormancy in bulblets of Lilium speciosum generated in vitro. I. The effects of culture conditions. Plant Cell Tiss. Org. Cult. 1990, 22, 167–172. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Boitel-Conti, M. The role of temperature in the growth and flowering of geophytes. Plants 2013, 2, 699–711. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Xie, Z.; Wu, Z.; Wang, Y.; Guo, Z.; Zhang, Y.; Wang, R. Response of oriental hybrid lily (Lilium oriental cv. Sorbonne) flowers to exogenous fluridone and abscisic acid application and responses of endogenous abscisic acid and gibberellic acid 3. HortScience 2015, 50, 559–564. [Google Scholar] [CrossRef]
- Langens-Gerrits, M.M.; Miller, W.B.; Croes, A.F.; de Klerk, G.J. Effect of low temperature on dormancy breaking and growth after planting in lily bulblets regenerated in vitro. Plant Growth Regul. 2003, 40, 267–275. [Google Scholar] [CrossRef]
- Miller, W.B.; Langhans, R.W. Carbohydrate changes of Easter lilies during growth in normal and reduced irradiance environments. J. Amer. Soc. Hort. Sci. 1989, 114, 310–315. [Google Scholar]
- Gandin, A.; Gutjahr, S.; Dizengremel, P.; Lapointe, L. Source-sink imbalance increases with growth temperature in the spring geophyte Erythronium americanum. J. Exp. Bot. 2011, 62, 3467–3479. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N. From leaf to crop: Quantifying Photosynthesis Responses of Two Flower Crops. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 24 June 2019. [Google Scholar]
- Lucidos, J.G.; Younis, A.; Lim, K.B. Determination of flower bud initiation in Oriental hybrid lilies ‘Siberia’ and ‘Sorbonne’. Pakistan, J. Agric. Sci. 2017, 54, 15–20. [Google Scholar]
- Almeida, D.B.D.; Barbosa, J.G.; Grossi, J.A.S.; Finger, F.L.; Heidemann, J.C. Influence of vernalization and bulb size on the production of lily cut flowers and lily bulbs. Semin. Ciências Agrárias Londrina 2017, 38 (Suppl. 1), 2399–2408. [Google Scholar] [CrossRef] [Green Version]
- Kohl, M.C., Jr. Correlation between rate of leaf initiation and diameter of Lilium longiflorum cultivar ‘Ace’. HortScience 1967, 2, 15–16. [Google Scholar]
- Roh, M.S.; Lawson, R.H. Propagation and transplant production technology of new floral crops. In Transplant Production Systems; Kurata, K., Kozai, T., Eds.; Kluwer Academic Publishers: Hoevelaken, The Netherlands, 1992; pp. 1–20. [Google Scholar]
- Arnold, W.N. The selection of sucrose as the translocate of higher plants. J. Theor. Biol. 1968, 21, 13–20. [Google Scholar] [CrossRef]
- Ranwala, A.P.; Miller, W.B. Sucrose-cleaving enzymes and carbohydrate pools in Lilium longiflorum floral organs. Physiol. Plant. 1998, 103, 541–550. [Google Scholar] [CrossRef]
Figure 1.
Shoot length measured at the end of each bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a), ‘Sorbonne’ (b), and ‘Siberia’ (c). *** Significant at p < 0.001.
Figure 1.
Shoot length measured at the end of each bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a), ‘Sorbonne’ (b), and ‘Siberia’ (c). *** Significant at p < 0.001.

Figure 2.
Contents of three soluble sugars and total sugar in bulbs measured at the end of each bulb storage periods at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d,g,j), ‘Sorbonne’ (b,e,h,k), and ‘Siberia’ (c,f,i,l). NS, *, **, *** Non-significant or significant at p < 0.05, 0.01, or 0.001, respectively.
Figure 2.
Contents of three soluble sugars and total sugar in bulbs measured at the end of each bulb storage periods at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d,g,j), ‘Sorbonne’ (b,e,h,k), and ‘Siberia’ (c,f,i,l). NS, *, **, *** Non-significant or significant at p < 0.05, 0.01, or 0.001, respectively.

Figure 3.
Plant height at the harvest of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a), ‘Sorbonne’ (b), and ‘Siberia’ (c). Within-graph means followed by the same letter are not significantly different by Tukey’s honestly significant difference test at p ≤ 0.05. NS, *** Non-significant or significant at p < 0.001, respectively.
Figure 3.
Plant height at the harvest of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a), ‘Sorbonne’ (b), and ‘Siberia’ (c). Within-graph means followed by the same letter are not significantly different by Tukey’s honestly significant difference test at p ≤ 0.05. NS, *** Non-significant or significant at p < 0.001, respectively.

Figure 4.
Fresh and dry weights of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d), ‘Sorbonne’ (b,e) and ‘Siberia’ (c,f). *, **, *** Significant at p < 0.05, 0.01, or 0.001, respectively.
Figure 4.
Fresh and dry weights of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d), ‘Sorbonne’ (b,e) and ‘Siberia’ (c,f). *, **, *** Significant at p < 0.05, 0.01, or 0.001, respectively.

Figure 5.
Correlation between days to flowering and plant height (a–c), fresh (d–f) and dry (g–i) weights of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d), ‘Sorbonne’ (b,e), and ‘Siberia’ (c,f). NS, *, **, *** Non-significant or significant at p < 0.05, 0.01, or 0.001, respectively.
Figure 5.
Correlation between days to flowering and plant height (a–c), fresh (d–f) and dry (g–i) weights of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’ (a,d), ‘Sorbonne’ (b,e), and ‘Siberia’ (c,f). NS, *, **, *** Non-significant or significant at p < 0.05, 0.01, or 0.001, respectively.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of leaves, leaf area, and SPAD index (chlorophyll content) at the harvest of cut flowers as affected by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’.
Table 1.
Number of leaves, leaf area, and SPAD index (chlorophyll content) at the harvest of cut flowers as affected by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’.
| Storage Period (days) | ‘Le Reve’ | ‘Sorbonne’ | ‘Siberia’ | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No. of Normal Leaves | Leaf Area (cm3) | SPAD Index | No. of Normal Leaves | Leaf Area (cm3) | SPAD Index | No. of Normal Leaves | Leaf Area (cm3) | SPAD Index | |
| 30 | 50.3 ab z | 608 d | 49.7 d | 28.0 b | 692 b | 49.2 ab | 27.9 ab | 1222 a | 61.5 ab |
| 40 | 43.3 ab | 483 e | 49.1 d | 31.0 ab | 810 ab | 52.8 ab | 29.8 a | 1192 ab | 62.0 ab |
| 50 | 50.3 ab | 853 a | 55.4 abc | 30.9 ab | 805 ab | 55.9 a | 24.3 b | 1046 ab | 60.5 ab |
| 60 | 51.9 a | 708 cd | 54.3 bc | 30.4 b | 746 b | 53.7 ab | 25.6 ab | 1100 ab | 60.0 ab |
| 70 | 46.4 ab | 713 bcd | 57.3 ab | 33.1 ab | 879 ab | 51.7 ab | 25.6 ab | 1100 ab | 60.0 ab |
| 80 | 47.4 ab | 817 ab | 59.9 a | 30.7 ab | 1030 a | 52.0 ab | 29.8 a | 1104 ab | 64.8 a |
| 90 | 49.0 ab | 766 abc | 57.7 ab | 36.4 a | 1033 a | 47.3 b | 25.9 ab | 1002 abc | 58.2 b |
| 100 | 48.0 ab | 696 cd | 55.3 bc | 29.3 b | 791 ab | 48.1 ab | 28.8 ab | 1012 ab | 59.7 ab |
| 110 | 46.8 ab | 622 d | 52.5 cd | 31.2 ab | 810 ab | 52.5 ab | 26.2 ab | 969 bc | 56.4 b |
| 120 | 41.7 b | 675 cd | 55.4 abc | 32.5 ab | 778 ab | 51.6 ab | 25.1 b | 790 c | 56.1 b |
z Within-column means followed by the same letter are not significantly different by Tukey’s honestly significant difference test at p ≤ 0.05.
Table 2.
Days to flowering, flower bud number, and flower diameter of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’.
Table 2.
Days to flowering, flower bud number, and flower diameter of cut flowers as influenced by bulb storage period at 0 °C in Lilium Oriental Hybrids ‘Le Reve’, ‘Sorbonne’, and ‘Siberia’.
| Storage Period (days) | ‘Le Reve’ | ‘Sorbonne’ | ‘Siberia’ | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Days to Flowering | No. of Flower Buds | Flower Diameter (cm) | Days to Flowering | No. of Flower Buds | Flower Diameter (cm) | Days to Flowering | No. of Flower Buds | Flower Diameter (cm) | |
| 30 | 80.6 a z | 2.7 a | 16.6 a | 105.8 a | 4.4 ab | 18.2 a | 120.2 a | 2.4 abc | 21.8 ab |
| 40 | 74.5 b | 2.3 ab | 16.8 a | 98.3 b | 5.0 a | 19.9 a | 112.9 b | 2.1 bc | 23.8 a |
| 50 | 71.0 c | 2.7 a | 17.1 a | 97.5 b | 4.0 ab | 20.4 a | 108.3 c | 2.8 ab | 21.8 ab |
| 60 | 70.9 c | 2.3 ab | 16.9 a | 91.6 c | 4.3 ab | 19.1 a | 106.3 c | 2.1 bc | 22.1 ab |
| 70 | 67.4 d | 1.8 ab | 15.1 b | 84.6 d | 4.4 ab | 18.3 a | 96.3 d | 2.1 bc | 22.1 ab |
| 80 | 62.7 e | 2.6 a | 15.4 b | 78.5 e | 3.9 ab | 18.7 a | 92.4 e | 2.8 ab | 22.3 ab |
| 90 | 62.3 e | 2.4 ab | 15.9 b | 74.8 ef | 4.3 ab | 16.1 a | 87.6 f | 2.3 abc | 20.7 b |
| 100 | 58.5 f | 2.3 ab | 15.6 b | 73.5 f | 3.5 ab | 17.1 a | 82.7 g | 3.1 a | 21.2 b |
| 110 | 53.9 g | 1.2 b | 15.2 b | 66.8 g | 3.7 ab | 18.1 a | 79.2 h | 2.3 abc | 20.3 b |
| 120 | 52.7 g | 1.3 b | 15.2 b | 66.5 g | 2.8 b | 18.3 a | 77.6 h | 1.6 c | 20.8 b |
z Within-column means followed by the same letter are not significantly different by Tukey’s honestly significant difference test at p ≤ 0.05.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, J.; Oh, W. Cold Storage Period of Bulbs Influences Their Sugar Contents and Post-Storage Growth and Flowering in Lilium Oriental Hybrids. Agriculture 2021, 11, 1080. https://doi.org/10.3390/agriculture11111080
AMA Style
Kim J, Oh W. Cold Storage Period of Bulbs Influences Their Sugar Contents and Post-Storage Growth and Flowering in Lilium Oriental Hybrids. Agriculture. 2021; 11(11):1080. https://doi.org/10.3390/agriculture11111080
Chicago/Turabian StyleKim, Jiseon, and Wook Oh. 2021. "Cold Storage Period of Bulbs Influences Their Sugar Contents and Post-Storage Growth and Flowering in Lilium Oriental Hybrids" Agriculture 11, no. 11: 1080. https://doi.org/10.3390/agriculture11111080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.